

مروری بر خواب بذر و مکانیسم ها وعوامل موثر بر آن
بررسی انواع خواب و جوانهزنی بذر
خواب و جوانه زنی : بذر گیاهان زراعی چگونه واکنش نشان میدهند؟
اینکه آیا بذرها جوانه میزنند یا مکانیسم خواب را انتخاب میکنند، از طریق فرآیندهای فیزیولوژیکی بسیار پیچیده اتخاذ میشود. زمان بندی صحیح این فرآیندها برای چرخه زندگی گیاهان بسیار مهم است. در صورت مواجهه با شرایط مرطوب، سطح خواب پایین باعث جوانه زنی قبل از برداشت در گونههای مختلف زراعی مانند گندم، ذرت و برنج میشود، این امر باعث کاهش عملکرد محصول و تأثیر منفی بر فرآوری پس از برداشت و صنایع تبدیلی میشود. در مقابل، سطح خواب عمیق بذر حتی در شرایط مطلوب از جوانه زنی طبیعی و استاندارد جلوگیری میکند و در نتیجه سرعت سبز شدن در مرحله کاشت و در نهایت فرآیند تولید پایین میآید. بنابراین، سطوحی از خواب بذر برای سیستمهای کشاورزی مکانیزه و مدرن الزامی است. طی چند سال گذشته، مطالعات متعددی نشان داده است که عوامل مختلف درونی و محیطی تعادل بین خواب و جوانه زنی را تنظیم میکنند، مانند نور، دما، وضعیت اقلیمی، میزان رطوبت و باکتریهای موجود در خاک، و فیتوهورمونهایی مانند ABA (اسید آبسیزیک) و GA (جیبرلیک اسید). در این مطالعه، پیشرفتهای اخیر در مورد مکانیسمهای مولکولی زمینهساز تنظیم فرآیندهای خواب و جوانه زنی بذر، از جمله فاکتورهای محیطی و هورمونی، بر گونههای زراعی مهم مورد بررسی قرار میگیرد .
بذر مهمترین نهاده کشاورزی و از طرفی نقطه آغاز و پایان چرخه زندگی بسیاری از گیاهان گلدار و در واقع واحد بنیادین تکوین حیات در زندگی گیاهان میباشد. استقرار موفق از لحاظ مکانی و زمانی و همچنین توان رویشی گیاهچههای جوان بطور عمده توسط حالتهای فیزیولوژیک و بیوشیمیایی بذر تعیین میشود. جوانه زنی بذرها وابسته به عوامل مختلفی از قبیل آب قابل دسترس، دمای مناسب، اکسیژن کافی، عدم وجود بازدارندهها و در برخی موارد کمیت و کیفیت نور است. با این وجود در برخی مواقع حتی با مساعد بودن شرایط محیطی و ژنتیکی بذر، در یک دوره مشخص از زمان، بذرها قادر به جوانه زنی نمیباشند که به چنین بذرهایی، بذرهای خفته گفته میشود. در شرایط خفتگی، بذرها زنده اما از نظر جوانه زنی غیرفعال هستند. خفتگی میتواند در بذر بسیاری از گونههای زراعی و وحشی وجود داشته باشد و مختص گونه خاصی نمیباشد. خفتگی میتواند به بذرها در زمانی که روی گیاه مادری قرار دارند القا شود که این نوع خفتگی سبب تأخیر یا عدم جوانه زنی میشود و خفتگی اولیه نام دارد.
انواع خفتگی بذر
- خفتگی ناشی از پوسته بذر
- خفتگی مورفولوژیک
- خفتگی فیزیولوژیک :
بذرهای دارای خفتگی فیزیولوژیک، جنین تمایز یافته و کاملاً توسعه یافته دارند ولی یک عامل بازدارنده فیزیولوژیکی از جوانه زنی ممانعت میکند. خفتگی فیزیولوژیک به علت حضور مواد بازدارنده یا نبود مواد تحریک کننده جوانه زنی یا ترکیبی از هر دو در بذر ایجاد میشود. مهمترین بازدارنده جوانه زنی در بذرها اسید آبسیزیک میباشد که بیشتر در پوسته بذر و در بافتهایی که با بذر در ارتباط مستقیم هستند وجود دارد.
- خفتگی مورفوفیزیولوژیک
- خفتگی چندگانه(ترکیبی)
در تیره گندمیان خفتگی فیزیولوژیک بیشترین سهم و کمترین سهم مربوط به خفتگی مورفولوژیک میباشد. موثرترین تیمارها جهت رفع خفتگی فیزیولوژیک استفاده از تیمارهای استراتیفیکاسیون سرد، جیبرلیک اسید و نیترات پتاسیم بود که با اعمال این نوع تیمارها تعادل هورمونی به سمت افزایش تحریک کنندههای جوانه زنی به نسبت بازدارنده جوانه زنی یعنی آبسیزیک اسید پیش میرود. هنگامی که بذرها از گیاهان مادری برداشت یا ریخته میشوند فرآیندهای فیزیولوژیک در بذرها درحال انجام است و وضعیت خفتگی بذرها بسته به محیط نگهداری دچار تغییرات قابل توجهی میشود.
بنابراین، سوال این است که چه چیزی تمایل بذر را برای جوانه زدن کنترل میکند؟ این شامل فرآیندهای پیچیده فیزیولوژیکی و بیوشیمیایی است که با سطح خواب بذر درون زا و نشانههای محیطی برون زا متنوع تعیین میشود (Gubler et al. 2005; Finkelstein و همکاران 2008).
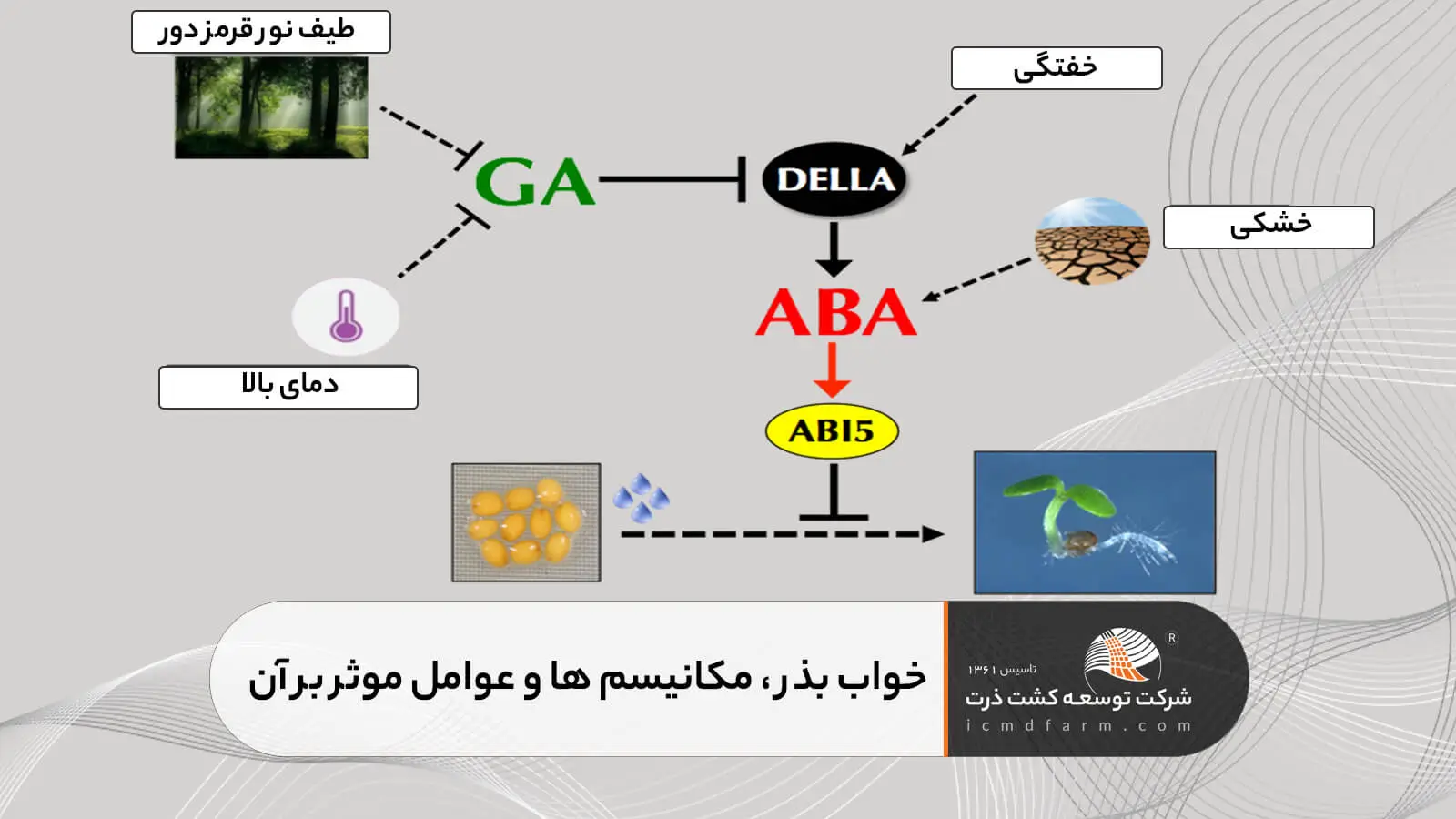
جوانه زنی قبل از برداشت و بر روی گیاه مادری یا مراحل انبارداری (PHS) در گونههای مختلف محصولات اصلی مانند گندم، ذرت و برنج، یکی از دلایل اصلی کاهش عملکرد و کیفیت محصول در سراسر جهان، به ویژه در مناطق مرطوب است (Fang et al. 2008; لیو و همکاران 2013). خواب بذر فرآیندهای جوانه زنی را حتی در شرایط جوانه زنی ایده آل به تأخیر میاندازد یا از آن جلوگیری میکند ، بنابراین از PHS نیز جلوگیری میکند. در آرابیدوپسیس مکانیسمهای مولکولی زمینه خواب و جوانه زنی بذر به طور فشرده و گسترده مستند شده است. مطالعات متعدد نشان داده است که عوامل زیادی خواب و جوانه زنی بذر را تنظیم میکنند و در این میان ، فیتوهورمونهای ABA (اسید آبسیزیک) و GA (اسید جیبرلیک) تنظیم کنندههای درون زا اولیه هستند که به طور متضاد خواب و جوانه زنی بذر را کنترل میکنند (Bewley 1997; Gubler و همکاران 2005; Finkelstein و همکاران 2008).
نکته مهم این است که هر دوی این هورمونها برای شرایط متنوع درونی گیاه و محرکهای محیطی که خواب و جوانه زنی بذر را تنظیم میکنند نقش کلیدی دارند، از جمله تولید اتیلن، BR (براسینوستریودها)، NO (اکسید نیتریک) و پاسخ به دما، کیفیت نور و غیره.
آبسیزیک اسید باعث خواب بذر میشود و مهار جوانه زنی میشود. در نتیجه، اکثر سیگنالهایی که باعث جهش میشوند یا هر سطحی از بیوسنتز ABA ، تغییر در سطوح خواب را ایجاد میکنند.
گندم که تقریبا غذای نیمی از مردم جهان میباشد، اولین گیاه زراعی اهلی شده است (گیل و همکاران 2004). برخلاف ذرت و برنج ، که بومی مناطق گرمسیری و نیمه گرمسیری میباشند ، گندم با مناطق دمایی متنوع سازگار شده است (گیل و همکاران 2004). بلوغ و رسیدگی سنبلههای گندم مشکل اساسی در تولید این محصول به ویژه در مناطق مرطوب است. علاوه بر کاهش عملکرد، PHS (جوانه زنی بر روی گیاه مادری یا مراحل انبارداری گندم) همچنین محتوای تغذیه ای دانه و کیفیت فرآوری را کاهش داده و در نهایت بر صنایع تبدیلی مربوطه اثر منفی میگذارد. جوانه زنی قبل از برداشت و بر روی گیاه مادری یا مراحل انبارداری گندم یک فنوتیپ پیچیده است در اثر برهمکنش بین ژنوتیپ و عوامل محیطی ایجاد و تنظیم میشود (امتیاز و همکاران 2008).
در گندم، رابطه جالبی بین رنگ دانه و PHS یافت شده است (Gfeller & Svejda 1960; Torada & Amano 2002). اگرچه مصرف کنندگان در بازار بین المللی دانه گندم سفید را به قرمز ترجیح میدهند، اما انواع گندم دانه سفید معمولا نسبت به نوع دانه قرمز به PHS حساس تر است (Groos et al. 2002; Himi و همکاران 2002). جوانه زدن قبل از برداشت در ذرت در اوایل سال 1930 گزارش شد (Mangelsdorf 1930). دانشمندان استنباط کردند که PHS ناشی از عدم حفظ خواب بذرها در مراحل رشد قبل از بلوغ است و علاوه بر این، این مکانیسم یک ویژگی ارثی است (Mangelsdorf 1930).
خواب بذر یک صفت بسیار پیچیده است که توسط عوامل مختلف درون زا و برون زا تعیین میشود. این موضوع اغلب به عنوان یکی از کمتر درک شده ترین پدیدهها در زیست شناسی بذر توصیف شده است (Bewley 1997; Finch-Savage & Leubner-Metzger 2006). از مطالعات قبلی، که در مورد گندم نشان داده شده است، یک مدل پیشنهاد شده که یک نمای کلی از مکانیسمهای تنظیمی زمینه خواب و جوانه زنی بذر ارائه میدهد. اگرچه پیشرفت سریعی از طریق تشریح ژنتیکی در گیاه مدل آرابیدوپسیس حاصل شده است، اما سوالات کلیدی متعددی در این زمینه همچنان باقی مانده است.
چه عواملی باعث کاهش ABA درون زا و افزایش GA در بذرهای پس از رسیدن میشود؟
ترکیبی از رویکردهای ژنومی و ژنتیکی باید برای بررسی فشرده رونویسی مکانی و زمانی خاص ژنهایی که به طور مستقیم یا غیرمستقیم بیوسنتز هر دوی این فیتوهورمونها را تنظیم میکنند، به کار گرفته شود. دادههای ژنومی و پس ژنومی درک ما را از تغییرات محتوای ABA و GA در طول بلوغ، ذخیره سازی و جوانه زنی بذر گسترش و عمیق تر میکند. مطالعات قبلی نشان داده است که برخی از مکانیسمهای تنظیم خواب بذر در میان گونههای مختلف زراعی حفظ شده است، به عنوان مثال ژنهای Vp1 در ذرت و آرابیدوپسیس (Koornneef et al. 1989; مک کارتی و همکاران 1989) و MFT در گندم و آرابیدوپسیس (Xi et al. 2010; ناکامورا و همکاران 2011; لیو و همکاران 2013). با این حال، تنوع ژنهای تنظیم کننده کلیدی هنوز در بین گونههای زراعی دست نیافتنی است. مسیر انتقال سیگنال ABA در محصولات زراعی نیز به وضوح درک نشده است ، اگرچه برخی از اجزا و عملکردها به طور مفصل مورد بررسی قرار گرفتهاند (کیم و همکاران 2012). گونه Zea mays L. به دلیل منشأ گرمسیری خود به دمای سرد حساس است. در عرضهای جغرافیایی بالاتر کشت که ذرت محصولی با اهمیت اقتصادی فزاینده است، گیاهچه ذرت در اثر بروز دمای کمتر از حدود 10 درجه سانتی گراد آسیب میبیند. فرضیهای که در دو مقاله (Capell and Dörffling 1993، Janowiak and Dörffling 1996) شکل گرفت و توسعه یافت، اخیرا توسط ریستیک و همکاران 1998 مورد بررسی قرار گرفت و دوباره با مواد و روشهای جدید مورد آزمایش قرار گرفت. بر اساس این فرضیه، ژنوتیپهای ذرت متحمل به سرما در مواجهه با تنش سرما، آبسیزیک اسید، سریع تر و در مقادیر بیشتری نسبت به ژنوتیپهای حساس به سرما تجمع مییابند. این رابطه مطابق با نقش محافظتی پیشنهادی ABA در برابر آسیب سرمازدگی است. همانند سایر تنش گیاهی، تصور میشود که هورمونهای گیاهی نقش کلیدی در پاسخ گیاه ذرت و سایر گونههای حساس به سرما به تنش سرما دارند. به نظر میرسد عملکرد محافظتی ABA ترکیبی است از: تثبیت غشاها، محافظت در برابر استرس اکسیداتیو (پراساد و همکاران 1994) تثبیت وضعیت آب با افزایش هدایت هیدرولیکی ریشه و بستن روزنهها (Markhart 1984، Eamus 1987، Ludewig et al. 1988)، القای ژنهای پاسخ دهنده ABA (اندرسون و همکاران 1994) و سنتز پروتئین (de novo) (Li 1994) تغییرات حضور کلسیم در مکانیابی سلولی و مکانیسمهای دیگر (لی و همکاران 1997). فرضیه به دو روش مورد آزمون قرار گرفت: اول با بررسی 20 ژنوتیپ جدید ذرت با تفاوتهای مشخص در تحمل سرما، دوم با دستکاری سطح درون زا ABA در طی تیمار دمای پایین با استفاده از بازدارنده بیوسنتز ABA، نورفلورازون، و اندازه گیری تحمل سرمای حاصل. تجمع ABA ناشی از دمای پایین در گروه ژنوتیپهای متحمل به سرما به طور معنی داری بیشتر از ژنوتیپهای حساس به سرما بود. این رابطه برعکس اتلاف آب ناشی از سرما بود. نتایج آزمایش دوم نشان داد که گیاهچههای تیمار شده با نورفلورازون نسبت به نهالهای تیمار نشده تحمل کمتری به سرما داشتند و ABA کمتری داشتند. کاربرد ABA کاهش تحمل سرما توسط نورفلورازون را جبران کرد. نتیجهگیری میشود که نتایج این فرضیه را تأیید میکند که تحمل سرما در ذرت با توانایی تجمع ABA به عنوان یک عامل محافظ در برابر آسیب سرمازدگی مرتبط است. میزان ABA در همه ژنوتیپها در طول سرمازدگی حتی در ژنوتیپهای حساس افزایش یافت، اما در ژنوتیپهای متحمل به میزان بیشتری افزایش یافت.
گردآورنده: مهندس پویان کاوسی
منابع





نظرات:
به منظور ثبت دیدگاه وارد سایت شوید!